|
RÙA HỒ GƯƠM, THẦN THÁNH HOÁ NHỮNG SAI LẦM KHOA HỌC
Bài viết lấy nguồn từ Tạp chí sinh học số 33(2): 30-39- 6-2011 - (Dịch theo nguyên bản tiếng Anh bài báo của Balázs Farkas, Lê Đức Minh và Nguyễn Quảng Trường “Rafetus vietnamensis Le, Le, Tran, Phan, Phan, Tran, Pham, Nguyen, Nong, Phan, Dinh, Truong and Ha, 2010 – another invalid name for an invalid species of softshell turtle (Reptilia: Testudines: Trionychidae)”. MỘT TÊN KHÔNG CÓ HIỆU LỰC KHÁC ĐẶT CHO MỘT LOÀI KHÔNG CÓ HIỆU LỰC THUỘC NHÓM RÙA MAI MỀM (LỚP BÒ SÁT REPTILIA: BỘ RÙA TESTUDINES: HỌ BA BA TRIONYCHIDAE) Russian Journal of Herpetology, 2011, 18(1): 65–72)
Tóm tắt: Bài này xem xét mô tả loài mới Rafetus vietnamensis của Lê Trần Bình và cộng sự công bố trong Tạp chí Công nghệ Sinh học năm 2010. Do tên loài được đặt cho cùng một mẫu vật chuẩn mà trước đây đã được mô tả là loài Rafetus leloii Ha (Hà Đình Đức, 2000), chúng tôi coi R. vietnamensis là tên đồng vật khách quan (objective synonym) của R. leloii. Đồng thời, do Lê Trần Bình và cộng sự không đưa ra được các đặc điểm xác đáng để có thể phân biệt được loài mới R. vietnamensi với loài đã biết R. swinhoei (Gray, 1873), nên chúng tôi khẳng định rằng hai loài này là một.
Từ khoá: Danh pháp, phân loại học, hình thái học, Rafetus vietnamensis, Rafetus leloii, Rafetus swinhoei, Việt Nam.
MỞ ĐẦU
Gần đây, Lê Trần Bình và cộng sự [cs.] (2010) đề xuất dùng tên Rafetus vietnamensis cho loài rùa mai mềm đã từng được công bố cách nay một thập kỷ bởi chính một đồng tác giả với tên loài là Rafetus leloii Ha (Hà Đình Đức, 2000), một loài không có sự khác biệt với loài Rafetus swinhoei (Gray, 1873) theo quan điểm của Farkas và Webb (2003) cũng như Lê Đức Minh và Pritchard (2009).
Công bố của Lê Trần Bình và cs. (2010) còn vi phạm các điều khoản và khuyến nghị ghi trong Luật Quốc tế về Danh pháp động vật (ICZN, 1999), – sau đây gọi tắt là Luật Danh pháp – ở một số điểm cũng như có nhiều chi tiết không nhất quán. Trong bài này, chúng tôi nêu lên những sai sót và do đó coi Rafetus vietnamensis là tên loài không có hiệu lực (invalid name).
Tên viết tắt của các cơ quan nghiên cứu viết theo Sabaj Pérez (2010), có bổ sung thêm các ký hiệu: BLF (Bộ mẫu vật của Balázs Farkas ở Gyúró, Hungary) và CPNP (Vườn Quốc gia Cúc Phương, Nho Quan, Ninh Bình, Việt Nam). Để tránh nhầm lẫn chúng tôi sử dụng tên viết tắt BMNH cho Bảo tàng Lịch sử tự nhiên (trước đây là Bảo tàng Lịch sử tự nhiên Anh), Luân Đôn, Vương quốc Anh và RMNH cho Trung tâm Đa dạng sinh học Hà Lan (trước đây là Bảo tàng Lịch sử tự nhiên), Leiden, Hà Lan.
VỀ MẶT DANH PHÁP
Theo Nguyên tắc Ưu tiên trong Luật Danh pháp (ICZN, 1999: Điều 23) thì “tên có hiệu lực của một đơn vị phân loại là tên hợp lệ (available name) được đặt sớm nhất”. Theo Farkas và Webb (2003), công bố mô tả loài Rafetus leloii của Hà Đình Đức (2000) đã đáp ứng được những yêu cầu tối thiểu của Luật Danh pháp như đưa ra mẫu chuẩn dùng để đặt tên (ICZN, 1999: Điều 16.4) và có mô tả ngắn, mặc dù còn thiếu sót về đặc điểm chẩn loại (ICZN, 1999: Điều 13.1.1), như vậy tên R. leloii Ha, 2000 đã có thể được coi là tên hợp lệ (mặc dù không có hiệu lực) và không cần thiết phải đặt tên mới để thay thế.
Theo Nguyên tắc về Mẫu chuẩn trong Luật Danh pháp (ICZN, 1999: Điều 61) thì “việc xác định mẫu chuẩn mang tên của một đơn vị phân loại hữu danh là tiêu chuẩn khách quan làm căn cứ cho việc áp dụng tên mà mẫu chuẩn đó mang”. Do R. leloii và R. vietnamensis có chung một bộ mẫu chuẩn (và mẫu vật tham khảo) nên chúng rõ ràng thuộc cùng một thực thể sinh học (ở đây được hiểu là cùng một loài). “Nếu một tác giả nào đề xuất một tên loài mới như một tên thay thế (…) cho một tên hợp lệ đã có từ trước, thì cả hai tên sẽ là tên đồng vật khách quan: cả hai đơn vị phân loại này có cùng một mẫu chuẩn mang tên, bất kể là tên mới thay thế chỉ được sử dụng hạn chế hoặc dùng cho các vật mẫu riêng biệt (ICZN, 1999: Điều 72.7). Từ việc sử dụng mẫu đồng chuẩn [allotype] của loài R. leloii làm mẫu chuẩn chính [holotype] cho loài R. vietnamensis và đưa mẫu chuẩn chính của loài R. leloii xuống vị trí các mẫu vật tham khảo của loài R. vietnamensis, Lê Trần Bình và cs. (2010: trang 950) – đã cố ý hoặc không cố ý – nhấn mạnh rằng họ coi R. vietnamensis là tên một loài mới. Tuy nhiên, “gợi ý” này sau đó lại mâu thuẫn với chính quan điểm họ đưa ra là “dữ liệu thu được từ tất cả các mẫu vật khác nhau ở Hà Nội, Thanh Hoá và Hoà Bình, đại diện cho sông Hồng, sông Mã và sông Đà, đều giống nhau và các mẫu vật giải khổng lồ thuộc nhóm Rùa mai mềm dã thấy ở miền Bắc Việt Nam đều thuộc cùng một loài duy nhất” (Lê Trần Bình và cs., 2010: trang 953; xem phần thảo luận của chúng tôi về mẫu vật NMW 30911 ở phần Kết quả phân tích sinh học phân tử). Bằng cách sử dụng cụm từ viết tắt “sp.nov.” (loài mới), Lê Trần Bình và cs. (2010: trang 950) một lần nữa chứng tỏ rằng các tác giả có dụng ý đặt tên cho một loài mới chứ không phải là đề xuất một tên thay thế (ICZN, 1999: Điều 16A). Mặt khác, các tác giả này (Lê Trần Bình và cs., 2010: trang 953) cũng cho rằng tên khoa học hợp lý nhất (…) phải là R. vietnamensis, còn tên loài R. leloii (bị viết nhầm là “R. leleoii”) “có liên quan nhiều đến truyền thuyết trả gươm ở hồ Hoàn Kiếm của vua Lê Lợi hơn là ý nghĩa về mặt phân loại học”, và điều này lại vi phạm một nguyên tắc khác của Luật Danh pháp, qui định rằng “tính hợp lệ (availability) của một tên không phụ thuộc vào tính không thích hợp (của tên đó)” (ICZN, 1999: Điều 18).
Điểm cần lưu ý khác nữa là Lê Trần Bình và cs. (2010: trang 950) ghi Rafetus leloii Ha, 2000 là loài chuẩn của giống Rafetus, thêm vào đó còn liệt kê thêm các tên khác như Rafetus hoankiemensis “VIR (tên viết tắt của báo Đầu tư) – vốn là một tên không có căn cứ (nomen nudum), và cả Rafetus vietnamensis “sp. nov” dưới cùng một tiêu đề này. Trên thực tế, loài chuẩn của giống Rafetus Gray, 1864 là Testudo euphratica Daudin, 1802 [nay đổi là Rafetus euphraticus] và được coi là loài duy nhất của giống này tại thời điểm đó (Gray, 1864: trang 81). Đáng lưu ý là tên họ Trionychidae bị ghi sai hai lần: đầu tiên là “Tryonychidae” (Lê Trần Bình và cs., 2010: trang 950) và sau đó là “Tryonichidae” (trang 952, chú giải cho Hình 5).
VỀ ĐẶC ĐIỂM CHẨN LOẠI
Theo Lê Trần Bình và cs. (2010: trang 950), các loài thuộc giống Rafetus khác biệt “rõ rệt với tất cả các giống khác ở các đặc điểm như (…) kích cỡ cơ thể, điều kiện sống, hình thái ngoài, hình thái sọ 33(2): 30-39 Tạp chí SINH HỌC 6-2011 và trình tự giải mã đoạn gen”. Theo quan điểm được các chuyên gia công nhận thì giống Rafetus hiện biết có 2 loài đang tồn tại, R. euphraticus và R. swinhoei (Meylan, 1917; Fritz & Havaš, 2007; Nhóm Chuyên gia Nghiên cứu về Phân loại rùa, 2007, 2009; Rhodin và cs., 2008). Rafetus euphraticus có chiều dài mai đạt tới tối đa 680 mm (Taskavak, 1998), như vậy rõ ràng phần lớn các loài thuộc các giống khác trong họ Ba ba (Tronychidae) như Amyda, Chitra, Nilssonia (theo Praschag và cs., 2007), Pelochelys và Trionyx có kích cỡ vượt qua con số này (tổng quan về kích cỡ của các loài thuộc họ Ba ba xem bài báo của Pritchard, 2001). Riêng với loài R. swinhoei chỉ tính các mẫu thu ở Trung Quốc thì tiêu bản mai lớn nhất có kích cỡ 860 mm (Lê Đức Minh và Pritchard, 2009). Tuy nhiên, với chiều dài mai (đo theo đường cong) là 1095 mm (theo Hà Đình Đức, 2000) thì mẫu vật ở Bảo tàng Hoà Bình mẫu chuẩn phụ (paratype) trong bài mô tả loài Rafetus leloii Ha, 2000 – vẫn nằm trong giới hạn về kích cỡ của các loài thuộc giống Chitra và Pelochelys (1000–1220 mm, xem Pritchard, 2001). Tất cả các mẫu vật khác của loài “Rafetus vietnamensis” đều có kích cỡ nhỏ hơn (xem Lê Đức Minh và Pritchard, 2009, với tên loài là R. swinhoei). Phải thừa nhận rằng những hồ nằm giữa khu vực đô thị như hồ Hoàn Kiếm không phải là sinh cảnh đặc trưng của bất kỳ loại ba ba nào. Tuy vậy, gần đây người ta cũng phát hiện một cá thể Cua đinh Amyda cartilaginea ở chính hồ này (theo ảnh chụp của Nguyễn Quảng Trường, [mẫu vật hiện đang lưu giữ tại Bảo tàng Động vật, Đại học Khoa học tự nhiên, Đại học Quốc gia Hà Nội: VNUH 2010.4.13]), và loài này cũng thường được nuôi trong các hồ gần đền thờ cũng như ở các ao hồ nhân tạo khác ở nhiều nơi trong khu vực Đông Nam Á. Thêm vào đó, sinh cảnh sống của loài “R.vietnamensis” bao gồm sông lớn và các đầm lầy – không phải là dạng sinh cảnh khác thường đối với các loài khác thuộc họ Ba ba.
Sự sai khác về hình thái ngoài và hình thái sọ cũng không rõ ràng – đáng tiếc là Lê Trần Bình và cs. (2010) đã không bình luận về vấn đề này. Bên cạnh đó, đặc điểm hình thái dễ nhận thấy nhất, đặc trưng cho giống Rafetus đó là sự tiêu giảm của đôi tấm sườn thứ 8 và sự phát triển của hai chai yếm không được Lê Trần Bình và cs. (2010) đề cập đến. Các kết luận và kết quả phân tích DNA (hay ADN) sẽ được thảo luận ở phần Kết quả phân tích sinh học phân tử.
VỀ MÔ TẢ ĐẶC ĐIỂM HÌNH THÁI
Mặc dù Lê Trần Bình và cs. (2010: trang 950) chỉ định mẫu vật đang lưu giữ tại chùa Hưng Ký (thuộc Bảo tàng Hà Nội), Hà Nội (số hiệu mẫu vật được nhóm tác giả này ký hiệu là HK1, không phải là số hiệu chính thức; Hà Đình Đức [2000] sử dụng ký hiệu là RHK 02-1968 cho chính mẫu vật này) là mẫu chuẩn chính của loài Rafetus vietnamensis, nhưng họ không đưa ra mô tả chi tiết về mẫu vật này. Mẫu vật này được minh hoạ bằng ảnh chụp mặt lưng có kích thước nhỏ và hình vẽ mô phỏng dựa trên cùng bức ảnh chụp mẫu vật (trang 951: Hình 1). Không có hình minh hoạ nào trong số này thể hiện “những sai sót khá kỳ quặc” về vị trí của một số tấm xương nhất định như Pritchard (2001) hay Lê Đức Minh và Pritchard (2009) đã đề cập đến, mà nhìn vào Hình 1 trong bài này, có thể thấy rõ hình vẽ này đã bị chỉnh sửa so với thực tế. Vết nối ở tấm sườn thứ 7 phía bên trái chắc chắn là một sản phẩm chế tác nhân tạo trong quá trình xử lý mẫu vật (xem hình 4A). Tuy vậy, Lê Trần Bình và cs. (2010) nhấn mạnh rằng bộ xương này “là đại diện tốt nhất cho tất cả các mẫu vật bởi sự nguyên vẹn của nó”. Nhóm tác giả chỉ đưa ra “những thông số về hình thái” (trang 951, ghi chú cho Hình 1) là kích cỡ xương mai (580 x 640 mm), kích cỡ xương sọ (150 x 236 mm) và tổng chiều dài bộ xương là 1686 mm. Cùng mẫu vật này, Hà Đình Đức (2000) cung cấp những số đo sau đây: dài xương mai 545 mm, rộng xương mai 510 mm, chiều dài cực đại xương sọ 236 mm, chiều rộng cực đại xương sọ 150 mm, và chiều dài gốc sọ 160 mm. Pritchard (2001) ước tính chiều dài xương mai khoảng 600 mm (được nhắc lại trong bài báo của Lê Đức Minh và Pritchard, 2009), trong khi đó Farkas và Webb (2003) dựa trên thước tỉ lệ trong bài của Hà Đình Đức (2000) ước tính chiều rộng sọ khoảng 160 mm. Hơn nữa, Lê Trần Bình và cs. (2010) chỉ đưa ra số liệu ước lượng của mẫu vật này và 2 mẫu khác được nhắc đến với ký hiệu là “NS1” cho mẫu thu ở hồ Hoàn Kiếm hiện đang trưng bày trong đền Ngọc Sơn, Hà Nội, mẫu này được Hà Đình Đức coi là mẫu chuẩn chính khi mô tả loài Rafetus leloii (“RHK 01-1967” trong bài của Hà Đình Đức [2000]; xem Hình 2) và “QL1” cho mẫu vật khác thu ở hồ Quỳnh Lâm gần thị xã Hoà Bình và hiện đang lưu giữ tại Bảo tàng Hoà Bình, mẫu chuẩn phụ của loài Rafetus leloii (“RHK 03-1993” trong bài báo của Hà Đình Đức [2000]; xương sọ được minh hoạ trong bài báo của Lê Trần Bình và cs. [2010: trang 951, Hình 3D] nhưng không chỉ rõ nơi lưu giữ mẫu sọ này), đồng thời số liệu này cũng không thống nhất với số liệu ghi trong phần mô tả: chiều dài cơ thể 1500–2000 mm, trọng lượng 170–220 kg (ở trang 950) so với 1700–2000 mm, trọng lượng 120 -220 kg (ở trang 951). Theo ước tính của Pritchard (2001) thì những mẫu này có chiều dài xương mai khoảng 635 mm (thảo luận về sự sai khác trong số đo của các tác giả khác nhau xem Farkas và Webb [2003: trang 109]), có thể thấy những mẫu này không thể có chiều dài cơ thể vượt quá 2000 mm.
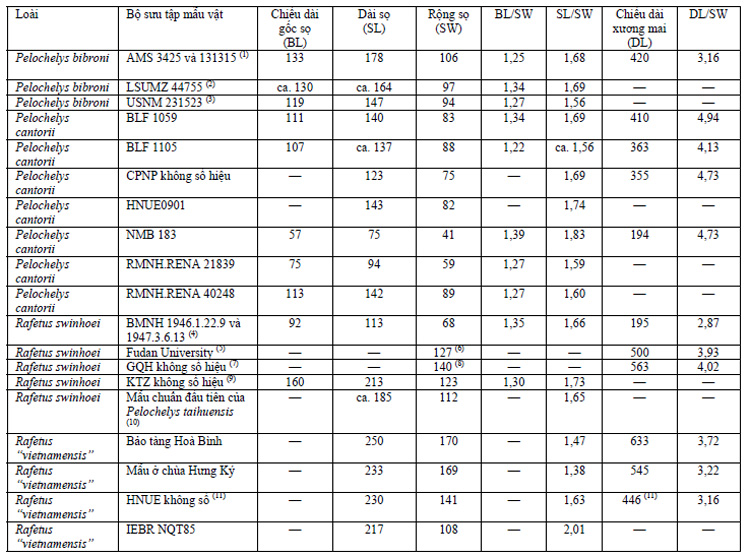
Cả phần đặc điểm chẩn loại và phần mô tả đều không đề cập đến bất kỳ đặc điểm hình thái nào phân biệt giữa loài R. vietnamensis và R. swinhoei. Đặc điểm khác biệt duy nhất mà Lê Trần Bình và cs. (2010) đưa ra so sánh là kích cỡ đầu: R. vietnamensis “có đầu to hơn và vòi mũi ngắn hơn so với Pelochelys bibroni và R. swinhoei”. Không có bất kỳ mẫu vật so sánh hoặc tài liệu tham khảo nào được đưa ra và luận điểm này của nhóm tác giả cũng không được chứng minh bởi các số liệu cụ thể. Hai giống Pelochelys và Rafetus khác biệt nhau ở một số đặc điểm (xem Meylan, 1987; Farkas và Fritz, 1998; Pritchard, 2001; Nguyễn Quảng Trường và cs., 2007), nhưng “kích cỡ đầu” không phải là một đặc điểm có thể dễ dàng so sánh về mặt định lượng. Mặc dù không có các mẫu vật tham khảo có kích cỡ tương đương trong các bảo tàng và trên thực tế các số liệu hiện có không thể so sánh được, nhưng căn cứ vào kết quả phân tích mẫu vật của chúng tôi thì xương sọ của loài Pelochelys cantorii phẳng hơn và hẹp hơn so với xương sọ của tất cả các loài thuộc giống Rafetus, ngoài ra tỉ lệ tương xứng giữa xương sau ổ mắt so với đường kính ổ mắt ở Pelochelys lại lớn hơn hẳn so với các loài Rafetus. Mặt khác, mặc dù tỉ lệ tương xứng giữa chiều rộng sọ với chiều dài gốc sọ hay chiều dài sọ có sự thay đổi rất lớn ngay trong từng đơn vị phân loại, tỉ lệ này lại không chênh lệch nhiều giữa các loài P. bibroni, P. cantorii, R. swinhoei hay “R. vietnamensis” (xem Bảng 1).
Để minh chứng luận điểm của mình, Lê Trần Bình và cs. (2010: Hình 3) trình bày một số ảnh xương sọ của giống Rafetus thu ở Trung Quốc và Việt Nam. Mặc dù không chỉ rõ mẫu vật thu ở Thượng Hải (Hình 3A) trong phần ghi chú nhưng có lẽ đây chính là sọ mẫu chuẩn chính của loài R.swinhoei (số hiệu BMNH 1947.3.6.13), đã được Nguyễn Quảng Trường và cs. (2007) so sánh và kết luận không có sự khác biệt so với mẫu sọ thu ở Bằng Tạ, Hà Tây (nay là Hà Nội) (ký hiệu mẫu IEBR NQT85) (Hình 3A trong bài báo này). Một mẫu sọ khác có nguồn gốc ở Quảng Phú, Thanh Hoá hiện đang lưu giữ tại Viện Công nghệ Sinh học, Hà Nội (Hình 3B trong bài này) thậm chí còn giống mẫu chuẩn BMNH 1947.3.6.13 hơn, chỉ trừ kích cỡ lớn hơn. Trên thực tế thì còn có sự khác biệt nhỏ đó là xương hàm trên của mẫu vật này (thu ở Thanh Hoá) tiếp xúc với nhau một phần trong khi đó ở mẫu chuẩn của loài R. swinhoei chúng lại tách biệt nhau bởi xương lá mía, tuy nhiên, đây chỉ là sự sai khác giữa các cá thể cùng loài như nhiều tác giả đã đã đề cập đến trong trường hợp loài R. euphraticus (xem Farkas và Fritz, 1998; Taskavak, 1999).Các đặc điểm chung khác của loài R. vietnamensis (hay của các loài thuộc họ Ba ba nói chung) được Lê Trần Bình và cs. (2010) nhắc đến như “đầu gắn với cổ dài, vòi mũi với hai lỗ rộng”, và “xương sọ rộng và tù” với “vòm hàm trên tù”. Hình 3 minh hoạ xương sọ mà nhóm tác giả này đưa ra cũng có dụng ý rằng “các mẫu ba ba này không thể thụt đầu vào trong mai được”. Hình 4 trong bài báo của chúng tôi cho thấy một số khác biệt đáng chú ý về hình thái xương mai ở các mẫu vật Rafetus của Việt Nam, ví dụ như tấm sống thứ 8 bị tách biệt ở mẫu chuẩn chính của loài R. vietnamensis (Hình A; mặc dù tấm sống thứ nhất và thứ hai dính liền nhau nhưng vẫn tính là 2 tấm theo cách đếm của Meylan [1987]). Sự tiêu giảm đôi tấm sườn thứ 8 chính là đặc điểm chẩn loại của giống Rafetus. Hướng sắp xếp các tấm sống bị đảo ngược từ tấm sống thứ 6, trong khi đó các đôi tấm sườn thứ 7 và thứ 8 tiếp xúc nhau với nhau theo một đường giữa lưng trên cả hai mẫu vật.
Một số “đặc điểm hình thái” khác của R. vietnamensis là “mai phẳng, thuôn dài, phủ lớp da mềm” và “da màu xanh xám đồng màu không có các vết đốm ở mặt trên” và “da uôn có màu hồng ở mặt dưới (yếm)” (Lê Trần Bình và cs., 2010: trang 950–951). Mặc dù sự thay đổi màu sắc ở các cá thể của loài R. swinhoei còn ít được đề cập đến trong các tài liệu, nhưng có thể thấy các vệt màu vàng trên đầu và mai ở cá thể non có thể chuyển thành đốm mờ hoặc biến mất hoàn toàn ở cá thể trưởng thành hoặc già (Pritchard, 2001; Farkas và Webb, 2003; Hình 5 trong bài báo này). Màu sắc của yếm cũng có thể thay đổi theo sự phát triển: các đốm mờ ở con non (Farkas, 1992; Vetter và van Dijk, 2006) sẽ biến mất theo độ tuổi, sau đó phần bụng trở nên có màu hồng hoặc màu vàng. Sự thay đổi về màu sắc theo độ tuổi rất phổ biến ở các loài ba ba.
|

|
|
| |
Hình 1. Bộ xương được lắp ghép không đúng của một cá thể rùa cái thu ở hồ Hoàn Kiếm, Hà Nội, được lưu giữ tại chùa Hưng Ký, Hà Nội, mẫu chuẩn chính của loài Rafetus vietnamensis Lê Trần Bình và cs., 2010. Ảnh: Nguyễn Quảng Trường.
|
Hình 2. Mẫu rùa mai mềm lớn thu ở hồ Hoàn Kiếm và được trưng bày trong đền Ngọc Sơn, Hà Nội, mẫu chuẩn chính của loài Rafetus leloii Hà Đình Đức, 2000. Ảnh: T. Ziegler. |
|
|
|
|
| |
Ghi chú: 1: Mẫu chuẩn mới (Neotype) của loài Pelochelys bibroni (Owen, 1853), mẫu tạo hình với xương sọ được tách riêng và có số hiệu riêng, số liệu theo Webb (1995). 2 và 3: Số liệu theo Webb (1995). 4: Mẫu chuẩn chính (holotype) của loài Rafetus swinhoei (Gray, 1873), mẫu ngâm với xương sọ đã được tách riêng, có số hiệu riêng; số đo dựa trên ảnh do hiện tại không thể đo trực tiếp trên mẫu xương sọ. 5: Không rõ số hiệu mẫu, số liệu theo Lê Đức Minh và Pritchard (2009). 6: Chiều rộng đầu của mẫu tạo hình. 7: Mẫu trong bộ sưu tập cá nhân của G. Q. Huang, Suzhou, Trung Quốc, số liệu theo Lê Đức Minh và Pritchard (2009). 8: Chiều rộng đầu của mẫu tạo hình. 9: Mẫu trong bộ sưu tập cá nhân của K. T. Zhao, Suzhou, Trung Quốc, số liệu theo Lê Đức Minh và Pritchard (2009). 10: Không rõ xuất xứ, số liệu theo Zhang (1984). 11: Theo Trần Kiên và Lê Nguyên Ngật, Hà Nội, mẫu xương sọ này và xương mai VNUH T91 rất có thể là từ cùng một mẫu vật.
|
|
|

|
|
| |
Hình 3. Mặt trên, mặt dưới và mặt bên xương sọ của các mẫu Giải Rafetus thu ở Bằng Tạ, Ba Vì, Hà Nội (IEBR NQT 85; A) và Quảng Phú, Thanh Hoá (Viện Công nghệ Sinh học, Hà Nội; B). Không có thước tỉ lệ. Lưu ý rằng xương hàm trên tách biệt nhau bởi xương lá mía ở hình A, trong khi đó chúng lại tiếp xúc nhau ở hình B. Ảnh: Nguyễn Quảng Trường.
|
|
|

|
|
| |
Hình 4. Xương mai mẫu chuẩn chính của loài Rafetus vietnamensis Lê Trần Bình và cs., 2010 (A) và VNUH T91 (Sông Mã, Thanh Hoá; B) với các khớp nối được tô rõ hơn. Vết nối trên tấm sống thứ 7 phía bên trái hình A được chế tác nhân tạo trong quá trình làm mẫu. Không có thước tỉ lệ. Ảnh: Nguyễn Quảng Trường.
|
|
|

|
|
| |
Hình 5. Thay đổi màu sắc ở trên đầu theo sự phát triển của loài Rafetus swinhoei: A, mẫu chuẩn chính của loài Oscaria swinhoei Gray, 1873 thu được tại Thượng Hải, Trung Quốc (số hiệu BMNH 1946.1.22. ảnh: Nguyễn Lân Hùng Sơn); B, mẫu vật ở Đồng Mô, Sơn Tây, Hà Nội, Việt Nam (Ảnh: T. McCormark); C, mẫu không rõ xuất xứ nhưng Niekisch và cs. (1997) cho rằng có nguồn gốc từ Việt Nam (ZMB 36437; ảnh: F. Hoehler, trong bộ sưu tập của U. Frizt); D, cá thể trưởng thành trước đây ở Vườn thú Thượng Hải, được cho là thu ở Gejiu, Vân Nam, Trung Quốc (theo Zhao và Adler, 1993; Lê Trần Bình và cs. [2010: trang 953] lại ghi là cá thể này được cho là “thu thập ở vùng thượng nguồn của sông Hồng của Việt Nam” và làm sai lệch thông tin về địa danh của Meylan [1987]; ảnh: J. Thorbjarnarson, trong bộ sưu tập của G. Schipper); E, đầu rùa rất lớn chụp ở hồ Hoàn Kiếm (ảnh trong bộ sưu tập của Nguyễn Quảng Trường). Không có thước tỉ lệ. Có thể thấy rõ sự thay đổi dần từ các đốm vàng rõ thành vân nhỏ và sau đó chuyển thành các đốm đen nhỏ trên nền da sáng màu.
|
|
VỀ KẾT QUẢ PHÂN TÍCH SINH HỌC PHÂN TỬ
Dựa vào kết quả phân tích sinh học phân tử, Lê Trần Bình và cs. (2010: trang 953) kết luận rằng “các mẫu giải khổng lồ ở miền Bắc Việt Nam là cùng một loài và duy nhất”, nhưng cùng lúc đó lại cho rằng chúng khác biệt với mẫu vật đang lưu giữ tại Bảo tàng Lịch sử tự nhiên Viên, Áo (số hiệu mẫu NMW 30911, [cũng được thu ở miền Bắc Việt Nam]), mà nhóm tác giả này đã công nhận là R.swinhoei. Tuy nhiên, ở phần này có một số vấn đề cần được làm rõ. Lê Trần Bình và cs. (2010) không mô tả chi tiết các quy trình chuẩn mà họ thực hiện trong quá trình nghiên cứu. Ví dụ như bài báo của các tác giả này không đưa ra thông tin cụ thể về quy trình tách chiết mẫu, trình tự của chuỗi mồi và điều kiện chạy PCR, như vậy rất khó để kiểm chứng kết quả của họ. Phương pháp sử dụng để phân tích mối quan hệ phát sinh loài cũng không được nêu rõ. Quan trọng hơn, kết quả giải mã gen của mẫu NMW 30911 chưa từng được công bố trên Ngân hàng Gen. Sau đó, chúng tôi đã cố gắng liên hệ với tác giả chính, Lê Trần Bình, để tham khảo kết quả giải mã gen nhưng cũng không thành công. Vì vậy, chúng tôi không thể xác nhận tính chính xác của số liệu mà nhóm tác giả này đưa ra. Hiện tại, chúng tôi cũng đang tiến hành phân tích lại DNA của chính mẫu vật này. Cũng cần thiết phải nhấn mạnh về khả năng sai sót trong quá trình phân tích sinh học phân tử của Lê Trần Bình và cs. (2010). Mẫu vật NMW 30911 có nguồn gốc thu ở miền Bắc Việt Nam, chứ không phải do Gray thu thập như ghi trong bài của Lê Trần Bình và cs. (2010: trang 953). Mẫu vật này được Farkas (1992) phát hiện lại và mô tả lại, cho thấy không có sự khác biệt đáng kể nào so với các mẫu vật khác của loài R. swinhoei. Nhưng căn cứ vào kết quả phân tích sinh học phân tử, Lê Trần Bình và cs. (2010) lại xếp nó có quan hệ gần gũi với giống Pelodiscus trong cùng nhánh tiến hoá Amydona theo Engstrom và cs. (2004), trong khi đó các mẫu vật khác thu được ở cùng vùng phân bố (miền Bắc Việt Nam) lại cùng nhóm với loài Rafetus euphraticus thuộc nhánh tiến hoá Apalonina theo Engstrom và cs. (2004). Kết quả này chứng tỏ giống Rafetus có hai nhánh tiến hoá khác biệt, và khả năng này rất khó xảy ra do các kết quả phân tích sinh học phân tử trước đây đều ủng hộ giả thuyết các loài thuộc giống này nằm trong cùng một nhánh tiến hoá (Lê Đức Minh và Pritchard, 2009). Bên cạnh những nhược điểm đã được Lê Đức Minh và Pritchard (2009) nêu ra có liên quan đến trình tự giải mã các đoạn gen cytb và ND4 (NADH4) được gửi đến Hệ thống Cơ sở Dữ liệu EMBL (Ngân hàng Gen), chúng tôi cũng thấy có một số sai sót và mâu thuẫn trong kết quả nghiên cứu của Lê Trần Bình và cs. (2010). Hình 4 chỉ là một bản đồ chỉ rõ các điểm ghi nhận phân bố của các loài “ba ba nước ngọt có kích cỡ khổng lồ ở Việt Nam”, nhưng sau đó Lê Trần Bình và cs. (2010: trang 952) lại chỉ dẫn Hình 4A–C tương ứng với kết quả về quan hệ phát sinh loài từ phân tích các gen 16S, cytb, và ND4. Thêm vào đó, mặc dù trong bài báo của các tác giả này chỉ nhắc đến 2 trình tự gen 16S được giải mã với số hiệu là AJ607405 và AJ607406 (cần lưu ý rằng các trình tự giải mã mà Lê Trần Bình và cộng sự công bố trên Ngân hàng Gen đều chỉ được ghi là “Rùa nước ngọt Việt Nam” mà không được định tên đến bậc giống hay bậc loài trên hệ thống cơ sở dữ liệu EMBL), cây phát sinh chủng loại (trang 952, Hình 5) chỉ dựa vào kết quả giải mã gen 16S và cho thấy có đến 3 trình tự của gen 16S của các mẫu Rafetus thu ở Việt Nam. Cũng theo cây phát sinh này thì giống Amyda cùng nhánh tiến hoá với các giống Aspideretes, Nilssonia, Dogania và Palea (trong khi đó theo Praschag và cs. [2007] thì giống Nilssonia trên thực tế bao gồm cả các loài thuộc giống Aspideretes), nhưng loài Apalone ferox thì tách biệt hẳn với Aplalone mutica và Apalone spinifera (ký hiệu viết tắt là “A. spiri” với “As. emoti” [viết đúng phải là A. spinifera emoryi?] tạo thành một nhánh tiến hoá riêng), kết quả này trái ngược hoàn toàn với kết quả phân tích về hình thái của Meylan (1987; tộc nhánh Apalonina) và Engstrom và cs. (2004, nhánh phát sinh Apalonina) cũng như kết quả phân tích sinh học phân tử của Lê Đức Minh và Pritchard (2009) vì Apalone và Rafetus phải ở chung cùng một nhánh tiến hoá. Các tác giả cũng không đưa ra giá trị thống kê gốc nhánh (bootstrap value) và cũng không giải thích vì sao lại chọn loài Cá sấu Caiman crocodilus (viết sai là “Caman crocodilus” ở cuối trang 952) làm đại diện cho nhánh phát sinh ngoài (cùng với loài Carretochelys insculpta).
KẾT LUẬN
Lê Trần Bình và cs. (2010) đã gây ra sự nhầm lẫn đáng tiếc trong mô tả loài Rafetus vietnamensis, trong đó các thông tin về phân loại học không hoàn chỉnh, không phù hợp hoặc có sai sót, các dữ liệu quan trọng hoặc còn thiếu hoặc bị thể hiện sai. Do cả hai tên loài Rafetus vietnamensis và Rafetus leloii được đặt ra cho cùng một thực thể sinh học, chúng tôi coi R. vietnamensis là một tên đồng vật khách quan của R. leloii. Cũng do Lê Trần Bình và cs. (2010) không đưa thêm được bất kỳ một chứng cứ mới nào để cho phép công nhận loài R. leloii, chúng tôi kết luận (lại) một lần nữa đây là tên đồng vật chủ quan (subjective synonym) của loài Rafetus swinhoei. Việc nghiên cứu kỹ hơn nhằm khẳng định sự phân hoá giữa các quần thể khác nhau trong phạm vi phân bố của loài này là cần thiết. Cuối cùng, việc so sánh giữa các mẫu vật của loài Giải thượng hải Rafetus swinhoei ở Việt Nam và Trung Quốc là rất quan trọng.
Lời cảm ơn. Chúng tôi rất biết ơn Heinz Grillitsch và Richard Gemel (Wien) đã gửi mẫu cơ của tiêu bản NMW 30911 để phân tích DNA, Ronald de Ruiter (Leiden) và Raffael Winkler (Basel) đã giúp chúng tôi đo đếm các mẫu vật. Cảm ơn Uwe Fritz (Dresden), Tim McCormark (Hà Nội), Nguyễn Lân Hùng Sơn (Hà Nội), Gerold Schipper (Frankfurt am Main) và Thomas Ziegler (Cologne) đã cung cấp ảnh, Mogens Andersen (Kobenhavn), Haral Artner (Sitzenberg-Reidling), Gábor Csorba (Budapest), Peter Paul van Dijk (Washington, D. C.), Wachira Kitimasak (Bangkok), Lê Nguyên Ngật (Hà Nội), Trần Kiên (Hà Nội) đã cung cấp ảnh và các thông tin có liên quan. Chúng tôi cũng đánh giá cao những ý kiến đóng góp của Wolfgang Boehme (Bonn) cho bản thảo của bài báo này.
TÀI LIỆU THAM KHẢO
Engstrom, T. N., Shaffer, H. B. and McCord, W. P. (2004), “Multiple data sets, high homoplasy, and the phylogeny of softshell turtles (Testudines: Trionychidae),” Systematic Biology, 53(5): 693 – 710.
Farkas, B. L. (1992), “Wiederentdeckung eines Exemplars von Rafetus swinhoei (Gray, 1873) im Naturhistorischen Museum Wien,” Salamandra, 28(2): 145 – 152.
Farkas, B. and Fritz, U. (1998), “On the identity of Rafetus swinhoei (Gray, 1873) and Pelochelys maculatus (Heude, 1880) (Reptilia: Testudines: Trionychidae),” Zoologische Abhandlungen Staatliches Museum für Tierkunde Dresden, 50(5): 59 – 75.
Farkas, B. and Webb, R. G. (2003), “Rafetus leloii Ha Dinh Duc, 2000 – an invalid species of softshell turtle from Hoan Kiem Lake, Hanoi, Vietnam (Reptilia, Testudines, Trionychidae),” Zoologische Abhandlungen, 53: 107 – 112.
Fritz, U. and Havaš, P. (2007), “Checklist of chelonians of the world,” Vertebrate Zoology, 57(2): 149 – 368.
Gray, J. E. (1864), “Revision of the species of Trionychidae found in Asia and Africa, with the descriptions of some new species,” Proceedings of the Zoological Society of London, 1864: 76 – 98.
ICZN (International Code of Zoological Nomenclature, 1999), 4th ed., Int. Trust Zool. Nomencl., London (available online at http://www.nhm.ac.uk/hosted-sites/iczn/code/).
Le, D. M and Pritchard, P. (2009), “Genetic variability of the critically endangered softshell turtle, Rafetus swinhoei: a preliminary report, pp. 84 – 92,” in: Ngo, D. C., Ta, H. T., Le, N. N., Hoang, X. Q., Vo, V. P., Nguyen, V. S., Nguyen, V. T., Le, T. T. and Tran, Q. D. (eds.), Proceedings of the 1st National Scientific Workshop on Amphibians and Reptiles in Vietnam, Hue University Press, Hue.
Le, T. B., Le, Q. H., Tran, M. L., Phan, T. H., Phan, M. T., Tran, T. T. H, Pham, T. T., Nguyen, D. T.,
Nong, V. H., Phan, V. C., Dinh, D. K., Truong, N. H. and Ha, D. D. (2010). “Comparative
morphological and DNA analysis of specimens of giant freshwater soft-shelled turtle in Vietnam related to Hoan Kiem turtle,” Tap chi Cong nghe Sinh hoc (Journal of Biotechnology), 8(3A): 949 – 954.
Nguyen, T. Q., Nguyen, V. S., Le, N. N. and Tran, K. (2007), “Identification of giant softshell turtle species (Reptilia: Testudines: Trionychidae: Pelochelys, Rafetus) in Vietnam,” Tap chi Sinh hoc (Journal of Biology), 29(2): 52 – 59 [in Vietnamese with an English abstract].
Niekisch, M., Farkas, B., Fritz, U. and Ha, D. D. (1997), “Rekordgrößen bei Weichschildkröten im Stadtzentrum von Hanoi, Vietnam,” herpetofauna, 19(107): 28 – 34.33(2): 30-39 Tạp chí SINH HỌC 6-2011
Praschag, P., Hundsdörfer, A. K., Reza, A. H. M. A. and Fritz, U. (2007), “Genetic evidence for wildliving Aspideretes nigricans and a molecular phylogeny of South Asian softshell turtles (Reptilia: Trionychidae: Aspideretes, Nilssonia),” Zoologica Scripta, 36: 301 – 310.
Pritchard, P. C. H. (2001), “Observations on body size, sympatry, and niche divergence in softshell turtles (Trionychidae),” Chelonian Conservation and Biology, 4(1): 5 – 27.
Rhodin, A. G. J., Van Dijk, P. P. and Parham, J. F. (2008), “Turtles of the world: annotated checklist of taxonomy and synonymy,” Chelonian Research Monographs, 5: 000.1 – 000.38.
Sabaj Pérez, M. H. (ed.) (2010), Standard symbolic codes for institutional resource collections in herpetology and ichthyology: an online reference, version 1.5 (4 Oct 2010), http://www.asih.org/.
Taskavak, E. (1998), “Comparative morphology of the Euphrates soft-shelled turtle, Rafetus euphraticus (Daudin, 1802) (Reptilia, Testudines) in southeastern Anatolia,” Amphibia-Reptilia, 19(3): 281 – 291.
Taskavak, E. (1999), “Cranial morphology of Rafetus euphraticus (Daudin, 1801) from Southeastern Anatolia,” Amphibia-Reptilia, 20(1): 35 – 53.
Turtle Taxonomy Working Group (2007), “An annotated list of modern turtle terminal taxa with comments on areas of taxonomic instability and recent change,” Chelonian Research Monographs, 4: 173 – 199.
Turtle Taxonomy Working Group (2009), “Turtles of the world: annotated checklist of taxonomy and synonymy, 2009 update, with conservation status summary,” Chelonian Research Monographs, 5: 000.39 – 000.84.
Vetter, H. and van Dijk, P. P. (2006), Turtles of the World, Vol. 4. East and South Asia, Edition Chimaira, Frankfurt am Main, 160 pp.
Webb, R. G. (1995), “Redescription and neotype designation of Pelochelys bibroni from southernNew Guinea (Testudines: Trionychidae),” Chelonian Conservation and Biology, 1(4): 301 – 310.
Zhang, M. (1984), “A new species of Pelochelys from Zhejiang, with subfossil description,” Acta Herpetologica Sinica, 3(4): 71 – 76 [in Chinese with an English abstract].
Zhao, E. and Adler, K. (1993), Herpetology of China, Contributions to Herpetology, No. 10, Society for the Study of Amphibians and Reptiles, Ithaca, 522 pp
|

